
Finch’s hypothesis – a brief history of escaping death
All organisms die, but not all organisms degrade at the same rate. In some species, death rates accelerate over a short period at advanced ages; others exhibit a slower, sustained loss in population numbers over the course of their lifespans. The term “senescence” describes these patterns as a function of physiological declines that occur with age. For decades, the general pattern of senescence – accelerated late-life mortality – was thought to be the standard rule of life. In the early 1990s, the universality of this thinking was called into question. In the 2000s, plant demographers motivated a more definitive case that patterns in birth and death rates were more diverse than previously recognized. A benchmark study by researchers at the Max Planck Institute for Demographic Research in 2014 drew upon the wealth of demographic studies to date to demonstrate a high level of diversity in the form of actuarial (i.e. mortality) and reproductive senescence – affirming the case that not all species exhibit positive senescence. We now know that some species do not decline at an accelerated rate with age (i.e. showing negligible senescence), and that some effectively age in reverse with respect to their demographic patterns (i.e. showing negative senescence). Demographers are actively seeking explanations for these unusual demographic patterns.
Caleb Finch posited a possible mechanism for these patterns in a monograph in 1990. This mechanism is based on the configuration of plant architecture. Unlike traditionally unitary organisms – where adult size is determined and physiological functions are highly integrated – plants can grow compartmentally, with indefinite growth and with low physiological integration across compartments. An individual plant, in other words, might be composed of many discrete functional units. Indeed, some ecophysiologists think of plants as a collection of straws that act more or less independently from each other. We call these units modules. Not all plants are modular, but modularity is largely a trait unique to plants and some animals, such as corals. Some have likened this physiological compartmentalization to physically coherent clones, individuals within an individual, or an organism with replicated organs. Thus, depending upon the threshold at which you assert functional integration as integral to how you define the individual, this question becomes partly a matter of scales of complexity and definitions of individuality. But exactly for this reason, the individual can die without dying.
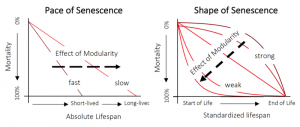
Under Finch’s hypothesis, death of the module is not closely aligned to death of the individual. Should injury be afflicted to one part of a modular organism, the injury may be kept local to that part of the organism. Loss in functionality will not render the physiological system dysfunctional for the entire organism – just the module. Injury will not scale. Thus, for a modular organism to die, it takes more than a local event that can cascade in its effect – it requires death by a thousand cuts compromising each of the individual modules.
In 2001, Douglas Larson argued that this trait, modularity, may be responsible for the extraordinary age that we see in relict populations of the coniferous tree northern white-cedar (Thuja occidentalis), and potentially bristlecone pine (Pinus longaeva). More recently, work has connected this trait with the susceptibility to air embolism. Air embolism describes the formation of an air bubble that impedes the continued conductance of water up the hydraulic column in plants. In some species, vessel architectures are modularized, and discrete. Consequently, these disruptive embolic events are isolated to the individual vessel or vessel-group. In other species, where lateral integration is high, these air pockets can quickly spread and prevent the movement of water and sap in the plant, causing death of the plant. The term “runaway embolism” is often used to describe these lethal events. For this reason, it was hypothesized that we would expect more modular architectures of plants in environments that are more conducive to the formation of air bubbles. Since emboli are partly a consequence of hydraulic stress, it was hypothesized that embolic risk is higher in more arid environments. Schenk and colleagues looked at this relationship in 2008 and discovered that there was a positive correlation between the degree of modularity and the degree of aridity, consistent with this line of thinking.
While the physical mechanisms and general landscape patterns align with evolutionary theory, there has not been a generalized test of Finch’s theory. We can not say, based on evidence to date, that modularity prevents the scaling of local injury to mortality and thereby increases life expectancy. Or for that matter, whether general evidence conforms with such a mechanism. To fill this crucial gap in our understanding of demography, anatomy, and evolutionary theories of senescence, we took advantage of existing datasets in demography and functional traits to test this question. Using functional traits that proxy modularity (e.g. intervessel distance, wood density), from TRY, we looked at whether the more sub-divided, compartmentalized anatomies really did promote longer life span across 138 plants from the COMPADRE database. We also looked at what this meant for the rate at which things die.
In the first comprehensive test of Finch’s hypothesis, we discovered evidence consistent with Finch’s prediction that modularity in plants correlates with longer life expectancy – as one would expect to see if modularity is preventing the scaling of local injury to injury of the individual. Our study also identifies for the first time, to the best of our knowledge, evidence that not just lifespan, but also age-dependent mortality are both correlated in part by the degree of modularity.
The next question was whether these relationships are Plant Kingdom-specific, or whether they are also found among animals. To explore this question, we tested whether organ multiplicity in 151 animals conformed similarly to the patterns we see in plants. Despite there being a high level of overall functional integration, we wanted to test whether there was a potential benefit of organ duplication on the redundancy of function. Could multiple organs enable system-wide function to be maintained after functional decline in another organ? We carefully examined the lymphatic and renal architectures in animals, and ranked their degree of multiplicity/subdivision on a hierarchical scale. In our test, we found a weak and conflicting signal. This would be consistent with the higher degree of physiological integration and key differences in the types of injury that tend to result in injury and death of the individual.
While far from the final word on this topic, our study sheds light on the influence of modularity. Out study suggests that modularity may shape lifespan and actuarial senescence rates in plants, but not in animals. Finch’s theory held that, just as a ship can float with a breached hull if the damage can be isolated to select compartments, plants and animals with modular anatomies would be more resilient to death if their anatomies prevent local injuries from becoming organismal in injury (i.e. fatal). Data advances have now allowed us to test Finch’s hypothesis across a wide range of plants and animals, and ask: Does a slowing down or even reverting of senescence correspond with more modular anatomies. Our review of 138 plants and 151 animal species suggests that Finch’s theory holds in plants, but possibly not in animals. Significantly lower rates of senescence are found in more modular plants, providing early evidence for a widespread phenomenon that lies at the heart of a crucial evolutionary puzzle. For more, please read our new manuscript, recently published in Functional Ecology.
Written by Connor Bernard

